グルコースの完全酸化と水との関係について、もう少し続けます。今回は、グルコースの完全酸化の代謝経路を、H2OとH+に着目して詳しく調べてみようと思います。便宜のために、二重標識水法(その1)の図2と3を再掲します。以下の説明に出てくる、数字を付した反応は、これらの図で示されています。
 |
図2
|
 |
| 図3 |
初めに、このブログを書くに当たって参考にした文献を掲げます。
15.Patel MS, Nemeria
NS, Furey W, Jordan F: The pyruvate dehydrogenase complexes: structure-based
function and regulation, J. Biol. Chem., 289, 16615-16623 (2014)
16. Ciszak EM, Korotchkina LG, Dominiak PM,
Sidhu S, Patel MS: Structural basis for flip-flop action of thiamin
pyrophosphate-dependent enzymes revealed by human pyruvate dehydrogenase, J.
Biol. Chem., 278, 21240-21246 (2003)
17. Wieser W: The splitting of water by the
tricarboxylic acid cycle Textbook error or textbook omission?, Trends Biochem.
Sci., 5, 284 (1980)
18. Madeira VMC: Stoichiometry of reducing
equivalents and splitting of water in the citric acid cycle, Biochem. Edu., 16,
94-96 (1988)
19. Hanson RW: Reducing equivalents in the
TCA cycle, Biochem. Edu., 19, 155 (1991)
1.水の関与する反応
グルコースの完全酸化の過程で、電子伝達系の前までに起きる反応(図2と図3)のバランス式を再掲します。
グルコース + 4 Pi + 2 ADP +2 GDP + 10 NAD+
+ 2 FAD + 2 H2O →
6 CO2 + 10 NADH + 2 FADH2 + 2 ATP + 2 GTP + 6 H+ ・・・反応式7
グルコース1分子当たり 2分子のH2Oが消費されます。図2と図3の反応でそのようになっているかを確かめてみます。H2Oが代謝中間体と反応するのは反応12と18で、計2分子のH2Oが使われます。ところが、反応9で1分子生成するので正味1分子の消費になります。グルコース1分子当たりでは2分子の消費となり、反応式7のH2Oの係数に一致します。
さらに、グルコース1分子当たり 4分子のPiが消費されることも次のように確認できます。Piと中間体との反応は、反応6と16で起き2分子のPiが消費されるので、グルコース1分子当たりでは4分子の消費となり、反応式7のPiの係数に一致します。次に、2つのH2Oが使われる反応と2つのPiの使われる反応を詳しく見てみましょう。
1) H2Oが使われる反応
H2Oと代謝中間体との反応は、グルコースの完全酸化の経路に2か所あます。反応12ではクエン酸合成酵素の反応の最後で、シトリルCoA が加水分解を受け、Oはクエン酸のカルボキシル基を構成し、一方のHはCoA側に入りチオールになり、他方のH はH+になります。反応18では、フマル酸の二重結合にHとOHとして取り込まれます。
 |
| 図4.グリセルアルデヒド 3-リン酸脱水素酵素の反応機構。-SHと-B:はそれぞれ活性中心にあるシステイン残基のチオール基とヒスチジン残基を表す。 |
2) Piの使われる反応
次に、Piの反応でO原子が中間体に取り込まれる反応6と16を詳しく見てみます。反応6を触媒するグリセルアルデヒド 3-リン酸脱水素酵素の反応機構を図4に示します。この酵素の活性部位にはシステイン残基があり、そのチオール基(-SH)が基質のアルデヒド基に結合してチオヘミアセタールを生成します。これがNAD+を還元するとアルデヒド基がカルボキシル基となってチオエステル結合が形成されます。Pi(HOPO32-)がこの結合のカルボニル(-C=O)を求核攻撃してアシルリン酸結合を形成し、生成物の1,3‐ビスホスホグリセリン酸が作られます。図2の反応6で示すように、1,3‐ビスホスホグリセリン酸のカルボキシル基のOの一つはPi由来です。また、H+がPiから遊離します。このような一連の化学変化の過程で、活性部位にあるヒスチジン残基(図4で‐B:と表示)がH+を受け取ったり受け渡したりして、触媒作用を発揮します。最後の段階で、ヒスチジン残基からH+が受け渡されてチオール基が再生されます。
図5.スクシニルCoA合成酵素の反応機構。https://commons.wikimedia.org/w/index.php?curid=15234158 を一部改変。
反応16はスクシニルCoA合成酵素によって触媒されます。その様子を図5に示します。初めに、PiがスクシニルCoAのチオエステル結合のカルボニルを求核攻撃し、CoASHの遊離をともなってスクシニルリン酸が生成します。続いて、活性部位のヒスチジン残基がスクシニルリン酸からリン酸基を受け取り、生成物のコハク酸ができます。最後に、ヒスチジン残基と結合したリン酸がGDPに移されてGTPが生成します。
反応6と反応16の反応メカニズムの比較から、基質レベルのリン酸化反応の化学的な戦略を見て取ることができます。両反応には次のような共通点があります。それは、①先ずチオールとカルボン酸の間でチオエステル結合が形成され、②次にPiがこの結合のカルボニル炭素を求核攻撃し、加リン酸分解を起こすことです。この分解によって、アシルリン酸ができてチオールが遊離します。③そして、アシルリン酸のリン酸をヌクレオシド二リン酸(ADPまたはGDP)が受け取ってヌクレオシド三リン酸(ATPまたはGTP)ができます。ここで登場するチオエステル、アシルリン酸、ヌクレオシド三リン酸はいずれもエネルギーに富む化合物で、この順にエネルギーが受け渡されてATPやGTPが生成します。
 |
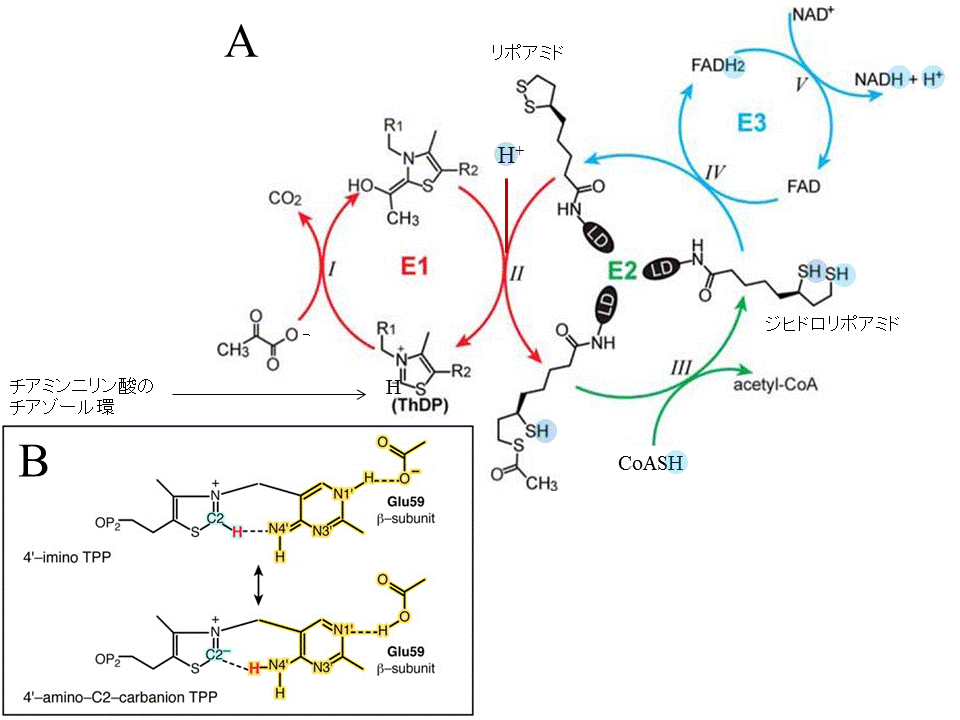
図6.ピルビン酸の酸化的脱炭酸反応。(A) ピルビン酸脱水素酵素複合体による酸化的脱炭酸の反応機構。文献15の図を一部改変。(B)ピルビン酸脱水素酵素(E1)の活性中心のチアミンニリン酸の構造。二つの構造が共鳴構造を取り、チアゾール環2位の水素が解離しやすくなっていることを示す。文献16の図を改変。
|
2.H+の収支
解糖系とTCAサイクルをつなぐ反応11(ピルビン酸の酸化的脱炭酸反応)は、H+との関係が分かりにくいので、初めにその概要を解説しておきます。図5Aに表すように、ピルビン酸脱水素酵素(E1)、ジヒドロリポアミド アセチル転移酵素(E2)、ジヒドロリポアミド脱水素酵素(E3)の3種類の酵素が順次働いて、ピルビン酸からアセチルCoAがつくられます。チアミンニリン酸を補酵素に持つE1が、ピルビン酸の脱炭酸(図のステップI)とE2に結合しているリポイル基の還元を行います。この還元の際、中間に生じるヒドロキシエチル基がリポイル基へ転移すると同時にアセチル基に酸化されます(ステップII)。E2によって、このようにして生成したアセチル基がCoASHに転移され、CoASH からのH+でジヒドロリポアミド基のチオールができます(ステップIII)。E3(FADを補酵素に持つ)が、その結果できたジヒドロリポイル基の電子2個がNAD+に渡されてNADH とH+を生成します(ステップIVとV)(文献15)。
ところで、反応11でH+が取り込まれるのは、上述のヒドロキシエチル基のリポイル基へ転移(ステップII)の際です。図4Aでは、反応前のE1のチアミンニリン酸のチアゾール環を、2位の炭素が水素を結合した形で表しました。X線構造解析の結果によれば、ヒトのE1の活性中心では、2位の炭素は水素が解離しやすい状態で結合している(図5B)と考えられるからです(文献16)。また、予測される反応機構によると、チアゾール環2位の水素は、H+となってヒドロキシエチル基のOH基に入ります(ステップI)。そのOH基の水素はヒドロキシエチル基がアセチル基に変わるときに、H+として離れてチアゾール環2位に移り、反応前のチアゾール環に戻ります(ステップII)。
電子伝達系で使われるNADH + H+を生成するうえで、H+の出入りがどのように起きるかを見てみます。図2(グルコースから2分子のピルビン酸ができる過程)と図3(ピルビン酸の酸化的脱炭酸とクエン酸サイクル)の全体の反応についてH+の収支を調べます。
1) グルコースから2分子のピルビン酸ができる過程
この過程を表す反応式を再掲します。
グルコース + 2 Pi + 2 ADP + 2 NAD+ →
2 ピルビン酸 + 2 ATP + 2 NADH + 2 H+ + 2 H2O・・・反応式4
以下の記述の末尾の括弧内の数は、グルコース1分子当たり経路から出るH+の個数です。マイナスは経路に入ることを示す。
○
反応1:基質がATPによってリン酸化されるとき、基質のOH基からH+が遊離する。(+1)
○
反応3:基質がATPによってリン酸化されるとき、基質のOH基からH+が遊離する。(+1)
○
反応6:加リン酸分解反応の所で説明したように、基質のアルデヒド基のHとPi のHを使ってNADH + H+が生じる。(+2)
○
反応10:ホスホエノールピルビン酸は、リン酸がADPに転移したあと、H+を取り込んでピルビン酸となる。(-2)
これらの反応の結果、収支は+2となり反応式4のH+の係数に一致します。
2)ピルビン酸の酸化的脱炭酸とクエン酸サイクル
この二つの過程を合わせると、ピルビン酸の完全酸化(電子伝達系の前まで)の反応になります。そのバランス式は(その1)の反応式5 と反応式6の各辺を足し合わせて、次のようになります。
ピルビン酸 + 4 NAD+ + FAD + GDP + Pi + 2 H2O →
3 CO2 + 4 NADH + FADH2
+ GTP + 2 H+・・・反応式10
以下の記述の末尾の括弧内の数は、ピルビン酸1分子当たり経路を出入りするH+の個数です。出るのをプラスとします。
○
反応11:ピルビン酸脱水素酵素複合体が触媒する酸化的脱炭酸反応で、補酵素チアミン二リン酸に結合したヒドロキシエチル基によってリポアミドが還元を伴ってアセチル化される際にH+が取り込まれる。(-1)
○
反応11:ピルビン酸脱水素酵素複合体が触媒する酸化的脱炭酸反応で、最終段階でジヒドロリポアミドが酸化されてNADH + H+が生じる。(+1)
○
反応12:シトリルCoAの加水分解によってCoASHとH+が生じる。(+1)
○
反応14:イソクエン酸の脱水素反応によってNADH + H+が生じる。(+1)
○
反応14:イソクエン酸は、脱水素反応のち脱炭酸が起きて生じた反応中間体に、H+が取り込まれてα-ケトグルタル酸が生成する。(-1)
○
反応15:α-ケトグルタル酸脱水素酵素複合体が触媒する酸化的脱炭酸反応で、補酵素チアミン二リン酸が触媒として働く際にH+が取り込まれる。(-1)
○
反応15:α-ケトグルタル酸脱水素酵素複合体が触媒する酸化的脱炭酸反応の最終段階で、ジヒドロリポアミドが酸化されてNADH + H+が生じる。(+1)
○
反応19:反応18でフマル酸にH2Oが付加されてできたリンゴ酸が、脱水素されてNADH
+ H+が生じる。(+1)
これらの反応全体では、収支は+2となり、反応式10のH+の係数に一致します。
3.CoASHの収支
以下の括弧内の数はピルビン酸1分子当たり出入りするCoASHの個数です。
○
反応11:アセチルCoAの生成過程でCoASHが使われる。(-1)
○
反応12:シトリルCoAの加水分解によってCoASHとH+が生じる。(+1)
○
反応15:スクシニルCoAの生成過程でCoASHが使われる。(-1)
○
反応16:スクシニルCoAの加リン酸分解によってCoASHが生じる。(+1)
これらの反応全体ではCoASHの収支は0で、反応式10と合います。
4.水の役割について
(その1)の記事で、グルコースの好気的酸化が、形式的に二つの反応(反応式2と3)に別けられることを述べました。反応式2を再度示します。
C6H12O6 + 6
H2O → 6 CO2 + 24 H+ +
24 e-
この式の6 H2O が6 CO2および 24 H+ + 24 e-の生成にどのように関わるかを代謝経路の反応と関連づけて見てみようと思います。
1)CO2のO原子の由来
CO2のO原子はグルコースの水酸基ないしアルデヒド基に由来するものと、H2OまたはPiに由来するものがあります。後者の場合、次のような過程でCO2に取り込まれます。
○
反応6: Pi(HOPO32-)のO原子1個が1,3‐ビスホスホグリセリン酸のカルボキシル基のOとなり、ピルビン酸のカルボキシル基のOとなります。ピルビン酸の酸化的脱炭酸反応(反応11)によってカルボキシル基からCO2ができます。
○
反応12: シトリルCoAの加水分解で使われるH2OのO原子は、CoAとチオエステル結合していたクエン酸のカルボキシル基に入ります。このカルボキシル基は、コハク酸のカルボキシル基となり、次のサイクルで代謝されるとCO2になります。
○
反応16: PiのO原子1個がスクシニルリン酸のリン酸と結合したカルボキシル基のOとなり、コハク酸を生成しますます。このカルボキシル基は、次のサイクルにおいてCO2になります。
○
反応18: H2OのO原子は、OH-としてフマル酸の二重結合に付加されてリンゴ酸のOH基となり、リンゴ酸が脱水素によってオキサロ酢酸のケト基になります。このケト基は、次のTCAサイクルでコハク酸のカルボキシル基になりますが、さらに次のサイクルにおいてCO2になります。
2) 24 H+ +
24 e-の由来
グルコースの完全酸化の過程で生成する5NADH+5H+ とFADH2の還元当量が何に由来するかを見てみましょう。5NADH+5H+を生成するのは、反応6,11,14,15および19です。そして、FADH2は反応17でできます。
○
反応6:図4に示すようにアルデヒド基からヒドリドイオン(:H-)がNAD+のニコチナミド環に転移します。H+はPiから生じます。
○
反応11:反応の中間でできるヒドロキシエチル基がリポアミドのジスルヒドに転移するとき2 e-を使ってジスルヒドを還元します。この2 e-はピルビン酸のカルボキシル基とケト基を結合していた電子対です。このときできるチオレートにH+が結合してジヒドロリポアミドの一方のチオール基をつくり、もう一方のチオール基は、アセチルジヒドロリポアミドからCoASHへアセチル基が転移するときにできます。最終的にジヒドロリポアミドがNAD+を還元してNADH+H+ を生成します。
○
反応14:イソクエン酸脱水素酵素によってイソクエン酸の第二級アルコールの炭素に結合した水素がヒドリドイオンとしてNAD+に転移してNADHが生成し、水酸基の水素はH+ になります。同時にアルコールはケト基に酸化されます。
○
反応15:反応11と類似の反応が進み、NADH+H+ を生成します。
○
反応17:-CH2-CH2- の一方の炭素のHがH+として、他方の炭素のHがヒドリドイオンとして引き抜かれFADに転移してFADH2を生じる。
○
反応19:リンゴ酸脱水素酵素によってリンゴ酸の第二級アルコールの炭素に結合した水素がヒドリドイオンとしてNAD+に転移してNADHが生成し、同時にアルコールはケト基に酸化され、H+ を遊離します。
5. 迷惑な論文
ここからは、蛇足かも知れません。話の切り出しに、生化学の教科書(「ヴォート基礎生化学」(東京化学同人)にある反応式を再掲します。
C6H12O6 + 6 H2O
→ 6 CO2 + 24 H+ + 24 e- ・・・反応式2
この式の両辺を比較すると、あたかもグルコースの12個のH原子と6H2Oの12個のH原子から24 H+ +
24 e-が生じたように見えます。H+の由来はそれでよいのですが、e-の由来はそうではありません。「ヴォート基礎生化学」には、この点について何もコメントされていませんが、反応式2の右辺の24 e-はすべてグルコースに由来し、H2Oは電子を供給しません。24 e-を与えるのは、グルコースのH:C結合とC:C結合の電子対です。形式的に次のように考えることができます。グルコースの好気的酸化では、C:C結合の電子対も代謝の過程で究極的にはH:C結合に転換され、合計12個のヒドリド(:H-)が脱水素反応によってNAD+やFADに転移されます。C:C結合の電子対がH:C結合に転換されるときにH2O に由来するH+ が使われます。
このようなことをわざわざ書いたのは、脳裏の片隅にあった或るジャーナルの記事を思いだしたからです。その記事は、タイトルが「The splitting of water by the tricarboxylic acid cycle. Textbook
error or textbook omission?」で、ミトコンドリア電子伝達系で使われる還元当量を作り出すのに、クエン酸サイクルによる水の開裂が必要であり、クエン酸サイクルが葉緑体の光合成と似た働きをすることを、一般生物学の教科書に載せようとしたが、編集者の反対にあったということを記しています(文献17)。この発想は、著名な生化学者Efraim Rackerが著書「A New
Look at Mechanisms in Bioenergetics」に書いたのが最初で、その後にも、同じ観点で書かれた論文がBiochemical Education誌に発表されました(文献18)。しかし、その後にこの論文を批判する記事が同誌にLetter to Editor として載りました(文献19)。さいわい標準的な生化学の教科書には、ミトコンドリアが水の開裂を行って還元当量を生成するというようなことは書かれていません。
化学が専門の知人(兵庫医科大学・福島和明教授)に、文献18の論点の正否について尋ねたところ、グルコースの完全酸化(C6H12O6 + 6O2 → 6 CO2+ 6 H2O)の両辺の各分子を構成する原子の「酸化数」を調べれば答えが得られるということでした。その説明の要点は以下の通りです。グルコースの完全酸化の場合、反応前後で、炭素原子の酸化数の合計は24増加し、酸素原子の酸化数の合計は24減少します。したがって、グルコースの炭素原子が合計で24 e-失い、その電子が全て酸素分子を構成する酸素原子に渡されることになり、この反応の電子のやりとりは完結します。
今回は、生体のグルコースの完全酸化にはH2Oの働きが重要であるので、代謝経路におけるH2OやH+の出入りにこだわってみました。瑣末なことにこだわり過ぎたかもしれませんが…。なお、化学の先生による、「酸化数」を使った酸化還元の説明は、私には目から鱗でした。手元の物理化学の本(齋藤勝裕著「絶対わかる物理化学」講談社)に、酸化還元の基本となるものに酸化数があり、酸化されるとは酸化数が増えることで、酸素(酸化数 -2)を受け入れるか、水素(酸化数
+1)もしくは電子を放出することであるという趣旨の説明がありました。長年生化学の教育に携わりながら、酸化数のことは不案内で忸怩たる思いがします。
無断転載禁止